El cuerpo humano puede, sin exagerar, figurar entre los sistemas biológicos más complejos del planeta. Nuestro cuerpo consta de miles de millones de células, muchos órganos y sistemas. Pero todo este esplendor y diversidad surgió debido a la fusión de solo dos células: un espermatozoide y un óvulo. Creo que no es necesario explicar cómo se realiza la fertilización y qué se requiere para esto (pista: las cigüeñas y el repollo no tienen nada que ver con esto). Pero aquí hay algunos aspectos de la vida de los espermatozoides que durante muchos años no estuvieron claros. Los científicos de la Universidad de Bristol, utilizando técnicas modernas de microscopía tridimensional, pudieron ver los movimientos de los espermatozoides de una manera que antes no era posible. Cómo y por qué medios se mueve el esperma, aprendemos del informe de los científicos. Vamos.
Base de investigación
A pesar de que los espermatozoides participaron en la creación de la vida mucho antes de la llegada del método científico, su camino en la literatura científica comenzó bastante recientemente, en 1677. El estudiante de medicina Johann Gam compartió sus observaciones con su colega y amigo Anthony van Leeuwenhoek (1632 - 1723), quien, a su vez, examinó y describió en detalle los "animales seminales" (como llamó a los espermatozoides).
Anthony van Leeuwenhoek / Lazzaro Spallanzani / Carl Ernst von Baer
Leeuwenhoek sugirieron que estas células inusuales están involucradas en la fertilización, pero su teoría, aunque cierta, fue rechazada por la comunidad científica. Durante mucho tiempo se creyó que los espermatozoides son parásitos y solo el fluido seminal está involucrado en la fertilización.
Solo casi cien años después, en 1786, Lazzaro Spallanzani (1729-1799) demostró el hecho de la participación de los espermatozoides en la fecundación en su obra Experiencias Para Servir a La Historia de La Generación De Animales y Plantas. Sin embargo, sus explicaciones para el proceso en sí eran bastante vagas: creía que el óvulo ya es el comienzo de un nuevo organismo y que el esperma solo es necesario para activar el proceso de crecimiento.
El mismo término "esperma" fue introducido a principios del siglo XIX por Karl Ernst von Baer (1792-1876).
Independientemente de lo que pensaran los científicos hace varios siglos, los espermatozoides tienen una función muy clara, cuya implementación es proporcionada por una serie de herramientas especializadas. La tarea principal del esperma humano es atravesar el tracto genital femenino, encontrar el óvulo y transferirle el material genético masculino.

La estructura del esperma La
célula reproductora masculina no puede presumir de dimensiones, porque es la más pequeña del cuerpo humano (excluyendo la cola): las dimensiones de la cabeza son 5.0x3.5x2.5 micrones (largo x ancho x alto), el largo de la parte media es 4.5 micrones, y longitud de la cola - 45 micrones.
Al mismo tiempo, el tamaño pequeño no es un inconveniente, sino un aspecto reflexivo de aumentar su velocidad. En el proceso de maduración de los espermatozoides, su núcleo (lleva un solo conjunto de cromosomas) se vuelve más denso, la mayor parte del citoplasma se descarta y solo los orgánulos más importantes permanecen en la célula.
El flagelo se puede llamar el segundo elemento más importante del esperma después del núcleo, es decir. su cola. Porque es gracias a él que se lleva a cabo el movimiento de esta célula a lo largo del tracto genital de una mujer. También es gracioso que el entorno vaginal sea extremadamente destructivo para las células germinales masculinas, pero el semen reduce parcialmente el efecto negativo sobre los espermatozoides. El nivel de pH dentro del tracto genital femenino permite que los espermatozoides se muevan hacia el útero, donde los espera un ambiente mucho más favorable.
Anteriormente, se creía que el esperma avanza debido al movimiento simétrico de su flagelo de lado a lado.
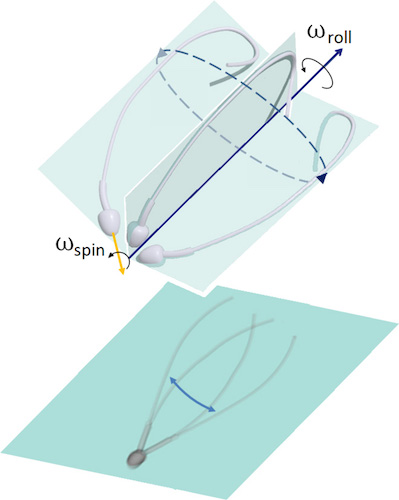
Imagen # 1: asimetría del movimiento flagelar en 3D (arriba); Proyección plana del movimiento del flagelo, creando una ilusión óptica de simetría bilateral en microscopía 2D (desde abajo).
Esta declaración se expresó incluso en los días de Levenguk. También condujo a una idealización simétrica de la forma de onda en tres dimensiones, a menudo percibida como una espiral cónica, similar a un sacacorchos en expansión.
Debido a la microscopía bidimensional, muchas observaciones se interpretaron de manera inexacta y, a veces, completamente incorrecta. Las afirmaciones sobre la simetría de los colgajos del flagelo contradicen muchas observaciones que muestran la asimetría estructural dentro del marco del propio flagelo.
Si el aleteo del flagelo de los espermatozoides durante el movimiento sigue siendo asimétrico, ¿cómo se logra la simetría del movimiento del flagelo de lado a lado y el movimiento de la célula hacia adelante? Es esta pregunta la principal en este estudio.
Para obtener una respuesta a esto, los científicos compararon observaciones moleculares y microscópicas que mostraban que el esperma humano usa tanto control asimétrico como anisotrópico para regular el aleteo flagelar. En otras palabras, la simetría se realiza debido a la asimetría: el efecto de una "parte superior de precesión" se produce cuando la cabeza gira ("fluido de perforación", palabras de los autores) simultáneamente e independientemente de cómo el flagelo del espermatozoide gira alrededor del eje de movimiento.
Resultados de la investigacion
El rápido movimiento de los flagelos de espermatozoides humanos se ha registrado con alta resolución espacio-temporal en 3D. Se examinaron dos grupos de espermatozoides que flotan libremente en un líquido de baja viscosidad: los espermatozoides que flotan junto al cubreobjetos (colocados en la parte superior de la muestra durante la microscopía) y lejos de él.
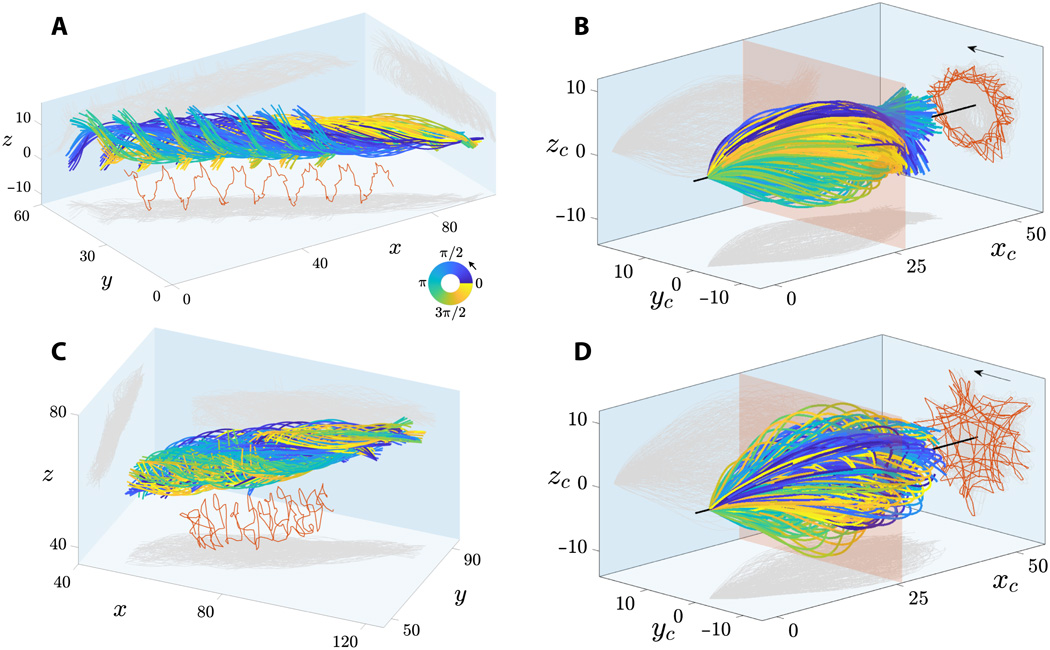
Imagen №2
En 2A y 2C se muestran flagelos de espermatozoides flotando cerca y lejos del cubreobjetos. Los colgajos de flagelo se caracterizan por un movimiento de rodadura característico alrededor de la dirección del movimiento de los espermatozoides.
La rotación combinada y el movimiento de traslación del flagelo del espermatozoide conduce a trayectorias en espiral del punto medio del flagelo con quiralidad preservada (marcado en rojo en 2Ay 2C ). Los espermatozoides exhiben rotación bidireccional alrededor de su eje: todas las células que flotan libremente (28 piezas) giraron en sentido antihorario cuando se ven desde el extremo frontal (flechas en 2B y 2D ) y solo 2 células giraron en el sentido de las agujas del reloj (no avanzaron debido a obstáculos en su camino).
Microscopía de espermatozoides flotando junto al cubreobjetos (corresponde a 2A ).
Todos los espermatozoides (30 células) se movió de manera similar a una tapa de precesión de rotación, en el que la rotación de la cabeza alrededor del eje longitudinal del espermatozoide (ω giro ) se produce simultáneamente y con independencia de cómo gira el flagelo con relación al eje de movimiento (ω rollo ).
En 2B y 2D se muestran los términos del flagelo aleteador del sistema de coordenadas concomitante * , es decir, un punto de vista que se mueve junto con el esperma, pero no un movimiento giratorio alrededor de su eje. Esto muestra que los golpes flagelares son extremadamente simétricos tanto en el plano ( xy ) como fuera del plano (z ) direcciones, que corresponde a observaciones en 3D.
El marco de referencia adjunto * es un marco de referencia asociado con el cuerpo en cuestión en un momento específico. El cuerpo dentro de este sistema está inmóvil. Por ejemplo, un ascensor que cae libremente es un marco de referencia que acompaña a un cuerpo que cae libremente en él, pero la Tierra no es tal sistema en relación con el cuerpo en un ascensor.La proyección de la trayectoria de la mitad del flagelo (líneas rojas en 2B y 2D ) muestra una increíble variedad de patrones geométricos, desde estrellas giratorias hasta triángulos, cuadrados y patrones de bucle con simetría polar. El patrón flagelar irregular que se muestra en 2D para los espermatozoides que flotan fuera del cubreobjetos también se observa para los espermatozoides que flotan junto al cubreobjetos. Esta variabilidad en los patrones puede ser causada por un desajuste en el desfase entre los componentes en el plano y fuera del plano del aleteo para cada celda, lo que aumenta en la línea de tiempo del movimiento rodante. En consecuencia, los patrones flagelares no son una característica distintiva de las células que flotan cerca y lejos del cubreobjetos.
Microscopía y modelado del flagelo que muestra la rotación de la cabeza y el flagelo alrededor del eje de movimiento.
La amplitud de la onda 3D se caracteriza por una curva envolvente simétrica en forma de bala, a diferencia de la espiral cónica (similar a un sacacorchos en expansión), que a menudo se describe en la literatura. Los espermatozoides que flotan lejos del cubreobjetos tenían una forma de onda más simétrica que las células que flotan junto al vidrio. Por tanto, el cubreobjetos adyacente es una fuente débil de asimetría para el colgajo flagelar.
Los espermatozoides que flotaban junto al cubreobjetos tenían un ángulo de ataque conservado * -7 °, con una orientación media del flagelo del esperma dirigida hacia el cubreobjetos.
* — ( ) , .

Imagen # 3
La imagen de arriba muestra una comparación de los golpes flagelares en el marco complementario (fila superior) y el marco rodante que lo acompaña (fila inferior).
La Figura 3E muestra la verdadera naturaleza del latido flagelar como se ve desde la perspectiva del esperma, sin ningún movimiento flotante o rodante.
El marco de referencia rodante adjunto ( 3E ) muestra que las aletas flagelares son anisotrópicas, es decir, Las características de las ondas en cada dirección transversal (perpendicular al plano de aleteo), etiquetadas como “ plano b ” (plano azul) y “ plano z ” (plano rojo), son marcadamente diferentes.
Si comparamos las áreas de color gris claro en los planos azul y rojo en 3E, entonces se puede ver que el aleteo es fuertemente asimétrico en el plano by se caracteriza por una simetría rota de izquierda a derecha, que recuerda a una forma de C.
Esta observación contrasta fuertemente con los patrones simétricos observados en el marco de referencia como móvil en 3A .
Forma de onda flagelar relativa a un marco de referencia fijo de laboratorio (x, y, z).
El análisis de componentes principales (PCA) de una forma de onda tridimensional ha permitido a los científicos descomponer la onda flagelar en varios "modos de forma básica", a los que aquí se hace referencia como modos PCA. La forma de onda 3D se puede reconstruir con buena precisión con solo dos modos de forma de onda, como se ve con 3B y 3F .
En 3C, se puede ver que los dos primeros modos de PCA son idénticos en forma, hasta una rotación de 90 °, capturando así la forma de espiral aerodinámica causada por el rodar de los espermatozoides.
En 3G , sin embargo, la forma de C asimétrica interna se detecta completamente solo por el primer modo PCA. Segundo modo PCA en 3G(resaltado en naranja) introduce pequeñas desviaciones perpendiculares al primer modo PCA (resaltado en azul), lo que indica que la forma de onda se puede descomponer en dos direcciones de onda transversales independientes que son de naturaleza anisotrópica.
El análisis de Fourier de los barridos permitió reconstruir el movimiento del flagelo utilizando solo dos modos de Fourier en 3D y 3H . En resumen, cada señal de oscilación común se puede aproximar mediante una simple suma de dos funciones:
f r (s, t) ≈ f 0 (s) + | f 1 (s) | pecado (ωt + φ (s))La primera función f 0 (s) no depende del tiempo, se llama "modo estático" y fija la asimetría de la señal promediada a lo largo de la longitud del arco (s) del flagelo.
La segunda función es una onda viajera sinusoidal, llamada "modo dinámico", que oscila a la frecuencia capturada por el primer pico del espectro de potencia de la señal.
La modulación de amplitud y fase de una onda viajera a lo largo del flagelo son iguales, respectivamente, | f 1 (s) | y φ (s) = arg (f 1 (s)).
Por tanto, la fase lleva información sobre las características de la onda viajera. Por ejemplo, si la fase φ (s) no cambia a lo largo de s, entonces la señal no es una onda viajera, sino una onda estacionaria.
Por lo tanto, el modo estático captura cualquier desajuste de forma de onda. En este caso, la línea recta negra en 3D refleja la simetría del barrido en los planos xy y xz en el marco comoving. Por el contrario, en el marco de referencia rodante adjunto ( 3H ), el modo estático (curva negra) se caracteriza por una gran amplitud asimétrica, que recuerda a una C.
Forma de onda del flagelo relativa al marco de referencia acompañante.
Forma de onda del flagelo en relación con el marco de referencia rodante que lo acompaña.
El modo dinámico en el marco complementario (curvas rojas) en 3D tiene una gran amplitud y es altamente simétrico en los planos xy y xz debido al movimiento de los espermatozoides.
Pero el modo dinámico en el marco de referencia rodante que lo acompaña (curvas rojas) en 3H tiene una amplitud de forma de onda reducida y una dirección de viaje preferida.
La reconstrucción de Fourier de la forma de onda se realizó sumando los modos estático y dinámico (gráficos en el centro en 3D y 3H ), que concuerda bien con las observaciones originales (gráficos a la derecha en 3Dy 3H ).
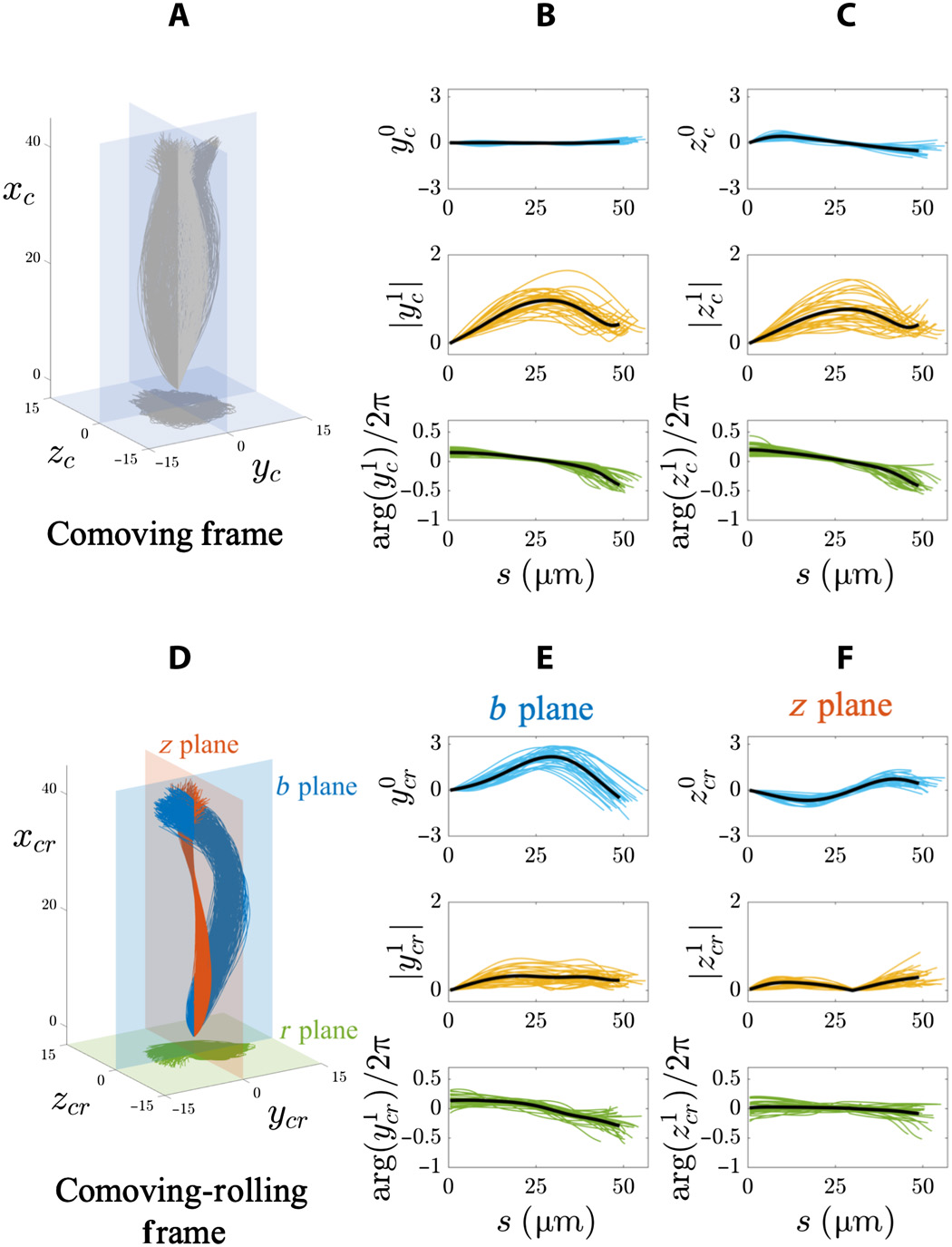
Imagen # 4 Los
gráficos de arriba muestran los resultados de un análisis de Fourier de un colgajo flagelar 3D en una población de espermatozoides que flotan libremente (20 células al lado del cubreobjetos y 8 células lejos del vidrio). En el marco de referencia adjunto ( 4A - 4C ), las amplitudes del modo estático en ambas direcciones (y cy z c ) son muy pequeñas (fila superior de gráficos en 4B y 4C ) debido a la simetría de los barridos.
Además, la amplitud (fila del medio) y la fase (fila inferior) de los modos dinámicos (y cy z c ) en 4B y 4Cfija la simetría transversal y excede la isotropía en este marco de referencia debido al hecho de que el espermatozoide gira alrededor del eje de movimiento.
Las características de la onda viajera para ambas coordenadas (y cy z c ) son las mismas para todas las celdas que flotan libremente: frecuencia 4 Hz, longitud de onda 100 μm y velocidad de onda 400 μm / s.
En el sistema de referencia de balanceo de acompañamiento ( 4D - 4F ), los regímenes estáticas distintas en la b y z planos demostrar que los columpios todavía tienen anisotropía en una población de células de libre flotación ( 4E y 4F ).
Modo estático b plano (y cr) es fuertemente asimétrico y sesgado hacia valores positivos (fila superior de gráficos por 4E ). Pero para el plano z (z cr ), oscila simétricamente a lo largo del arco de manera sinusoidal (fila superior en 4F ).
La amplitud de la régimen dinámico en el b plano (y 1 cr ) aumenta hasta alcanzar una meseta (fila del medio en 4E ), mientras que el régimen dinámico en el z plano (z 1 cr ) es no monótona a lo largo de la longitud del arco (fila del medio a 4F ). Características de las ondas viajeras y cr ( bplano) fueron: frecuencia 8 Hz, longitud de onda 145 μm y velocidad de onda 1120 μm / s. Características de una onda viajera z cr ( plano z ): frecuencia 6 Hz, longitud de onda 1526 μm y velocidad 5174 μm / s.
Los pequeños cambios de fase a grandes distancias a lo largo de toda la longitud del flagelo requieren velocidades de propagación de ondas muy altas. En consecuencia, las oscilaciones del plano z se comportan realmente como una onda estacionaria que pulsa en el tiempo.
Microscopía de espermatozoides no propulsores y rodantes.
La totalidad de los datos anteriores sugiere que el aleteo de flagelos utiliza dos controles transversales anisotrópicos coactivos, que no son muy diferentes de las ondas electromagnéticas viajeras. Sin embargo, cada onda de corte (y cr , z cr ) es la suma de los modos estático y dinámico: una onda viajera asimétrica a lo largo del plano b (azul en 4D ) y una onda estacionaria simétrica en el plano z (rojo en 4D ).
Es curioso (pero no sorprendente) que la presencia de un cubreobjetos cerca de la trayectoria del espermatozoide también tenga su efecto sobre la amplitud de la propagación de ondas.

Imagen No. 5 El
vidrio reduce la amplitud de la propagación de ondas debido a las interacciones hidrodinámicas entre el flagelo y la superficie dura del cubreobjetos.
En el marco complementario ( 5A y 5B ), la amplitud de ambos modos dinámicos (y c , z c ) disminuye hacia el final del flagelo espermático cerca del cubreobjetos (gráficos en el centro), mientras que los modos estáticos permanecen sin cambios (gráficos parte superior).
El modo dinámico z c es solo ligeramente menor que y c (curvas azules) en las gráficas centrales 5A y 5B . Esto contrasta con los perfiles simétricos e invariables de ambos (yc , z c ) modos dinámicos para espermatozoides detectados lejos del vidrio (curvas rojas en los gráficos del centro 5A y 5B ).
Si prestamos atención al sistema de referencia de rodadura que lo acompaña, entonces la naturaleza del efecto de vidrio es bastante anisotrópica, ya que afecta solo a un plano de rodadura, es decir, en el plano b ( 5C ).
El cubreobjetos afecta los modos y cr estáticos y dinámicos (gráfico superior y en el centro a 5C ). Pero el plano z (z cr ) permanece sin cambios (gráfico en la parte superior y central en 5D ).
La forma del modo estático en el sistema de referencia rodante complementario es la misma entre todos los espermatozoides y define una espiral derecha descentrada indicada por h (s) y mostrada por la curva negra en 5E . La proyección de la espiral sobre el plano rodante (plano verde) es una espiral en sentido antihorario que no tiene simetría polar, es decir, desplazamiento hacia un lado (proyección gris en 5E ).
Las espirales en modo estático se parecen mucho a las espirales logarítmicas que a menudo se encuentran en la naturaleza. Sin embargo, en este caso, el radio de la espiral cambia de manera no monotónica, aumentando / disminuyendo a un ritmo más rápido que el de las espirales logarítmicas en otras partes de la naturaleza.
Espiral logarítmica derecha h (s) en 5Ese puede expresar en términos de su radio y tono ( 5G ), que decae exponencialmente a lo largo del flagelo. Cualquier cambio en el signo h y o h z hace que la espiral cambie a la rotación en sentido horario, creando así una espiral a la izquierda. Todos los espermatozoides flotantes generaron espirales en sentido antihorario. Solo dos espermatozoides tenían espirales en sentido horario, aunque con una forma idéntica a 5E . Eran estas dos células las que no podían nadar hacia adelante debido a algunos obstáculos en su camino, pero podían continuar girando alrededor de su propio eje de torsión.
En todos los casos, la rotación de la hélice se correlaciona con la dirección de los espermatozoides rodando de la siguiente manera: una hélice en sentido antihorario para la rotación en el sentido de las agujas del reloj (visto desde la parte trasera) y una hélice en el sentido de las agujas del reloj para la rotación en sentido antihorario.
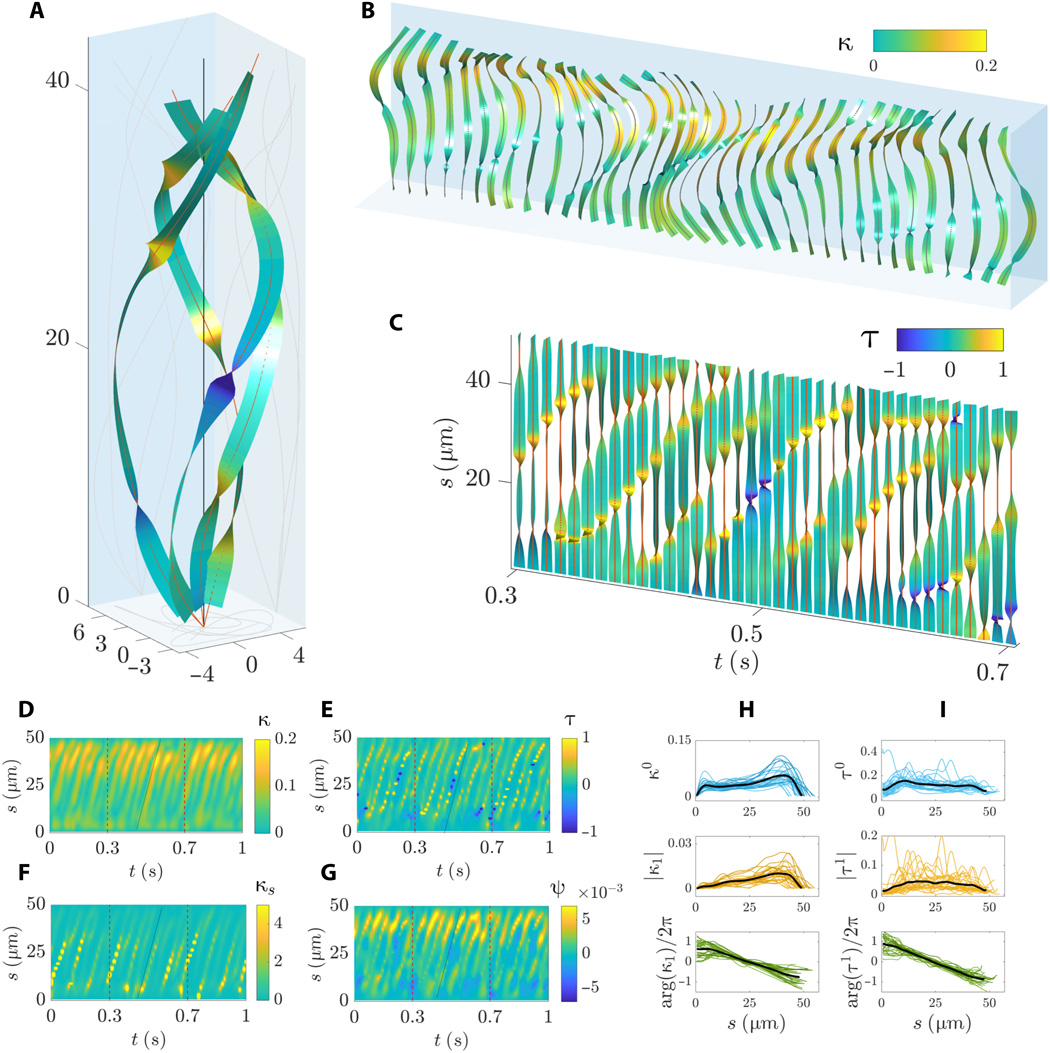
Imagen # 6 Los
gráficos 6A y 6B muestran una secuencia compleja de ondas viajeras a medida que el flagelo gira alrededor de su eje de rodadura. Las ondas de flexión se propagan linealmente a lo largo del flagelo con amplitud no monótona a lo largo del arco, caracterizadas por un fuerte aumento en las regiones media y distal.
La torsión de la forma de onda se caracteriza por curvas pronunciadas a lo largo de la longitud del arco ( 6B y 6C) con giros positivos y negativos simultáneos. La forma de espiral de la línea central del flagelo sufre un fenómeno por el cual coexisten regiones de quiralidad opuesta a lo largo del flagelo. Sin embargo, las partes del flagelo con quiralidad opuesta se mueven durante el aleteo ( 6C y 6E ). Las ondas de torsión viajeras se propagan a la misma velocidad que la onda de curvatura.
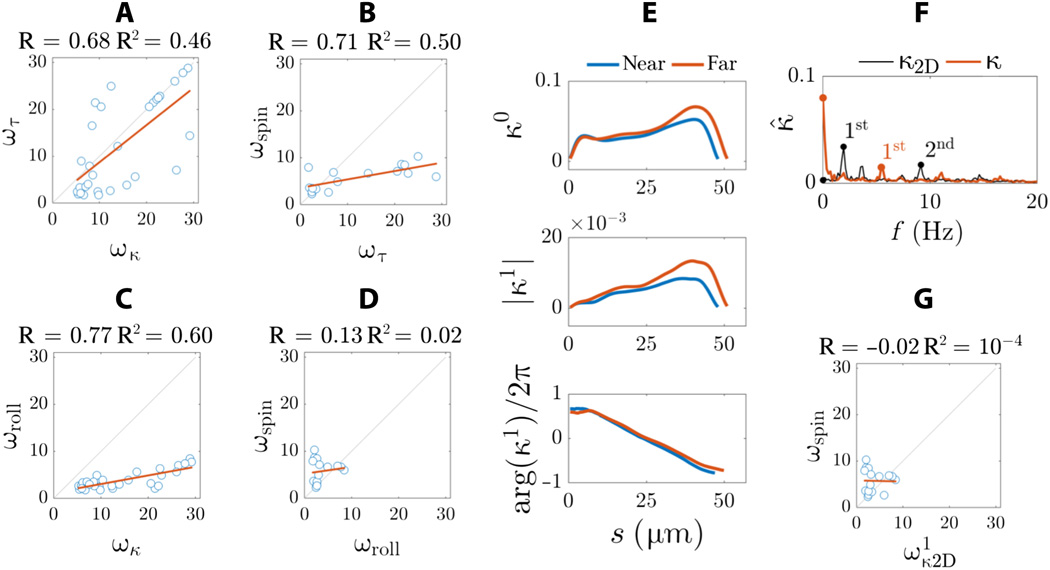
Imagen # 7
Para comprender mejor la diferencia entre los resultados de microscopía 3D obtenidos en este estudio y los resultados de microscopía 2D que a menudo se describen en la literatura, los científicos crearon una proyección 2D a partir de una forma de onda 3D.
El régimen estático de curvatura bidimensional es muy insignificante ( 7F). En 2D, la asimetría interna de la forma de onda no se puede detectar. En cambio, el espectro de frecuencia se caracteriza por dos picos de frecuencia (marcadores negros en 7F ) en lugar de un pico de frecuencia principal visto para la curvatura tridimensional (curva roja en 7F ).
Para un conocimiento más detallado de los matices del estudio, le recomiendo que consulte el informe de los científicos y materiales adicionales .
Epílogo
En este trabajo, los científicos pudieron demostrar en la práctica que, debido a la microscopía tridimensional, es posible descubrir lo que la microscopía bidimensional no dará, por ejemplo, para ver cambios en la dirección de la espiral (cambios en la quiralidad) durante la oscilación de los flagelos.
A primera vista, puede parecer que los espermatozoides simplemente flotan hacia adelante, moviendo la cola. Sin embargo, un examen detallado de la cinemática de estas células mostró que este proceso es mucho más complicado. El aleteo y balanceo de los flagelos, la rotación de la propia célula, todo esto junto permite que los espermatozoides avancen.
Parte de esta información estaba disponible anteriormente, pero no todos los detalles se han detallado debido a las limitaciones de la microscopía 2D. Comprender cómo se mueven los espermatozoides puede ayudar mucho en el campo de la medicina reproductiva, dicen los científicos. En particular, los nuevos datos ampliarán la gama de parámetros mediante los cuales se determinan las células sexuales no saludables de los hombres, que es la causa de la infertilidad en aproximadamente la mitad de las parejas.
En cualquier caso, cuanto mejor comprendamos ciertos fenómenos y procesos, especialmente los que tienen lugar en nuestro propio cuerpo, más probabilidades hay de que mejoren la calidad de nuestra vida.
Gracias por su atención, tengan curiosidad y tengan una buena semana de trabajo, chicos. :)
Un poco de publicidad
Gracias por estar con nosotros. ¿Te gustan nuestros artículos? ¿Quieres ver contenido más interesante? Apóyanos haciendo un pedido o recomendando a amigos, VPS en la nube para desarrolladores desde $ 4.99 , un análogo único de los servidores de nivel de entrada que hemos inventado para usted: Toda la verdad sobre VPS (KVM) E5-2697 v3 (6 núcleos) 10GB DDR4 480GB SSD 1Gbps desde $ 19 o cómo dividir el servidor correctamente? (opciones disponibles con RAID1 y RAID10, hasta 24 núcleos y hasta 40GB DDR4).
¿Dell R730xd es 2 veces más barato en el centro de datos Equinix Tier IV en Ámsterdam? ¡Solo tenemos 2 x Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 TV desde $ 199 en los Países Bajos!Dell R420 - 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB - ¡Desde $ 99! Lea sobre Cómo construir la infraestructura de bldg. clase con servidores Dell R730xd E5-2650 v4 a un costo de 9000 euros por un centavo?