
La evolución nos ha dado muchos animales inusuales. Algunos de ellos son tan inusuales que sus descubridores fueron llamados ladrones (como fue el caso de los ornitorrincos en 1797). Pero no solo la fauna puede presumir de especies no estándar, entre los representantes de la flora también hay cuervos blancos, es decir, plantas que no quieren sentarse en una dieta soleada, sino que prefieren insectos jugosos y otros animales pequeños como plato del día. Anteriormente, ya nos familiarizamos con un estudio en el que los científicos examinaron la trampa para moscas de Venus y sus espinas marginales.... Hoy tenemos que averiguar qué tan sensibles son estas espinas. Científicos de la Universidad de Zúrich (Suiza) llevaron a cabo una serie de experimentos prácticos, cuyo propósito era medir la velocidad de reacción del atrapamoscas venus al toque de una fuerza particular. ¿Cómo exactamente hicieron cosquillas los científicos al papamoscas, con qué rapidez reaccionó y cómo estos resultados explican las preferencias gastronómicas de la planta depredadora? Encontraremos respuestas a estas preguntas en el informe de los científicos. Vamos.
Base de investigación
Para no estirar esta narrativa, la descripción de la Venus atrapamoscas (que ya se usó en el artículo anterior) está oculta debajo del spoiler:
¿Qué es un papamoscas?
, . , . — , . , «» (, ) ( , ). 630 , , .
: . ( ), — , .

. , — ( 300 000 1 ) , . , , . , . , .
, «».
, . : , , . , , . , , , . -, . .

: , - .
, -, . . , . , . , . , ? , . 2 20 . , , . , , , , .
. , . , , . , ( ), , . . , , . — , . , . , 2-3 .
. , , . , , . , .
True Facts: Carnivorous Plants ( : ). .
, -. , , .
, , , , « ». , , «». , , , , . , . , .
: . ( ), — , .
. , — ( 300 000 1 ) , . , , . , . , .
, «».
, . : , , . , , . , , , . -, . .
: , - .
, -, . . , . , . , . , ? , . 2 20 . , , . , , , , .
. , . , , . , ( ), , . . , , . — , . , . , 2-3 .
. , , . , , . , .
True Facts: Carnivorous Plants ( : ). .
, -. , , .
, , , , « ». , , «». , , , , . , . , .
La idea principal del mecanismo de activación de la trampa del atrapamoscas Venus es que la futura presa necesita hacer dos toques en los sensibles pelos de la planta en 30 segundos. Cada toque de ellos produce un potencial de acción * necesario para cerrar la trampa. Los científicos han creído esto durante 200 años, pero ¿la teoría es consistente con la práctica?
A lo largo de los años de investigación, los científicos han desarrollado un escenario de eventos que deberían (en teoría) desarrollarse durante la caza de un papamoscas:
- una planta hambrienta atrae a los insectos secretando compuestos volátiles;
- insecto poco inteligente explora la planta (esperando comida gratis), durante la cual toca uno de los seis pelos sensibles, causando así un potencial de acción (AP por potencial de acción);
- 30 , «It’s a trap!», , ;
- (, ), (C12H18O3) .
Como resultado, los "pétalos" previamente abiertos del papamoscas se convierten en una especie de estómago, donde se digieren las presas fallidas.
En este trabajo, los científicos decidieron considerar en detalle el proceso de conversión de la estimulación mecánica de los cabellos sensibles en una señal eléctrica. Sabemos con certeza que tocar los cabellos sensibles abre canales iónicos mecanosensibles, pero todavía no hemos podido identificar estos canales.
Mientras estos canales putativos están abiertos, el potencial receptor * (RP procedente del potencial receptor ) aumenta, y si la desviación del cabello es lo suficientemente grande, RP alcanza un umbral por encima del cual se manifiesta AP.
* — , .
* — , .
* — .
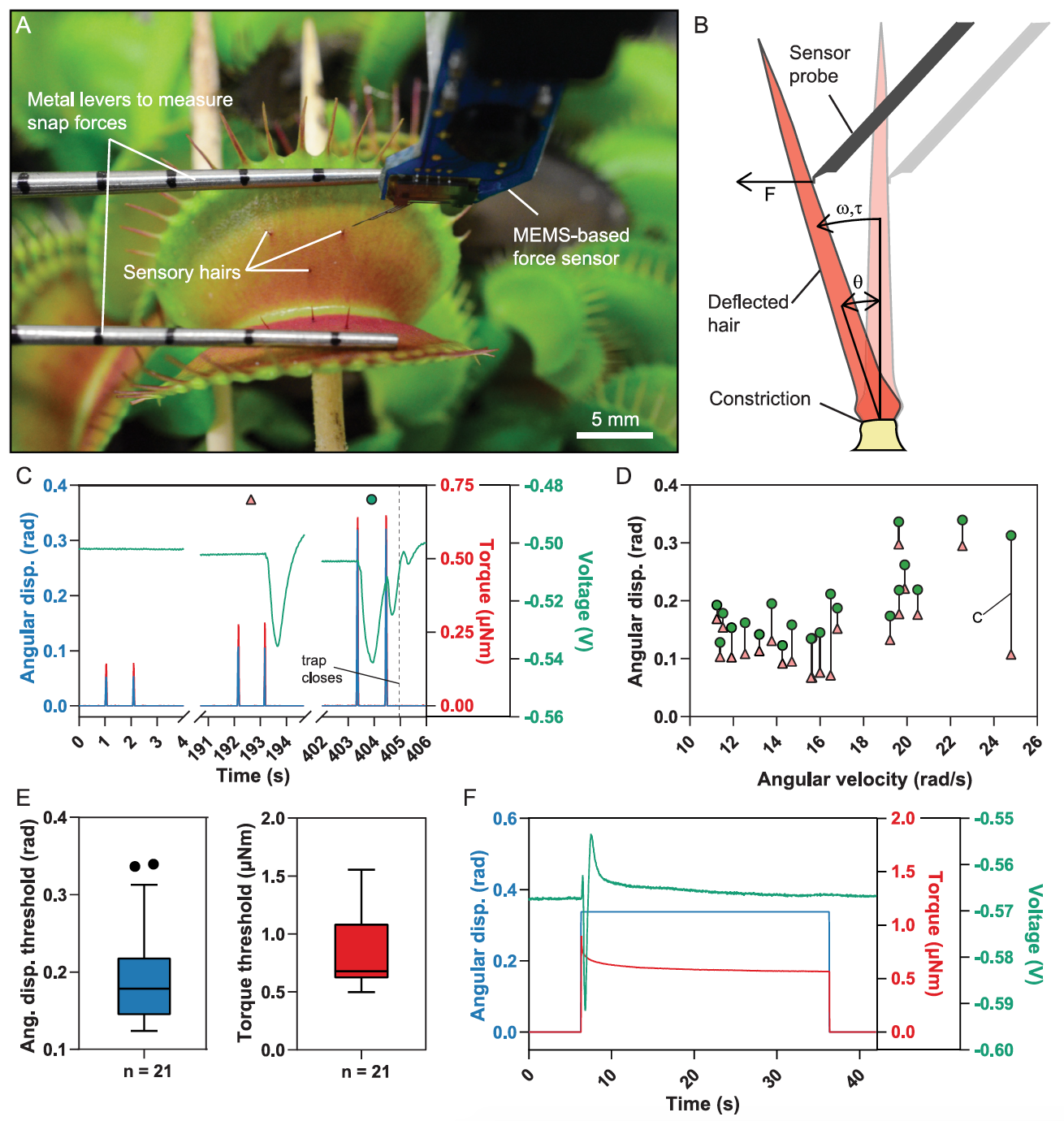
Imagen # 1
Ha habido intentos anteriores de vincular los estímulos mecánicos y la generación de potencial de acción, pero no había instrumentos adecuados para mediciones precisas. En el caso del estudio que estamos revisando hoy, los científicos utilizaron un sensor de fuerza basado en MEMS * montado en un sistema microrobótico para controlar con precisión la velocidad y amplitud de la deflexión y medir simultáneamente la fuerza aplicada ( 1A y 1B ).
Los MEMS * (sistemas microelectromecánicos) son dispositivos que combinan componentes microelectrónicos y micromecánicos.
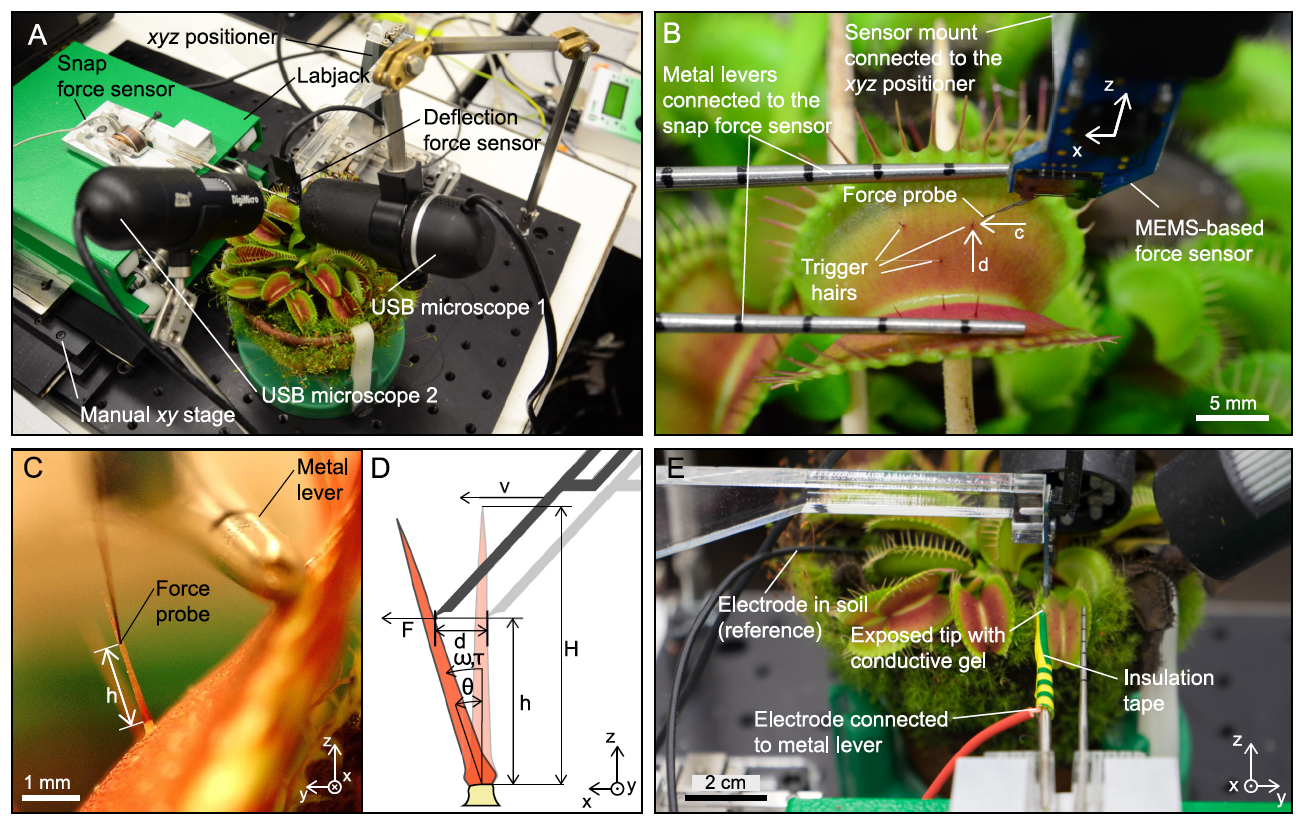
Imagen # 2
De esta manera, los científicos pudieron determinar con precisión el rango de parámetros en los que la desviación de los pelos conduce al cierre de la trampa, mientras que el segundo sensor de fuerza midió la fuerza instantánea generada (imagen de arriba).
Resultados de la investigacion
Durante los experimentos, los científicos tuvieron en cuenta que las arañas, moscas y hormigas (presas comunes del papamoscas) harán que los pelos sensibles se desvíen con bastante rapidez. En consecuencia, el sistema microrobótico se implementó a toda velocidad para imitar estos estímulos. Esto resultó en una alta velocidad angular inicial que variaba de 10 a 20 rad / s.
Estudios anteriores han encontrado que las hormigas desvían el cabello sensible a una velocidad angular de 0,25 a 7,8 rad / s, pero más lento que las moscas. Por lo tanto, los parámetros utilizados son más altos que los reales, lo que es muy adecuado para evaluar las capacidades del atrapamoscas Venus.
A velocidades tan altas, la duración de la deflexión es significativamente más corta que la de otros factores dependientes del tiempo involucrados, como la descomposición del RP y la relajación sensorial del cabello (1F ).
Tener en cuenta la desviación angular más que lineal permitió corregir las diferencias en la altura del contacto de la sonda sensorial con respecto al lugar de estrechamiento del cabello sensorial, así como las diferencias en cuanto a diferentes geometrías del cabello sensorial ( 1B , 2C y 2D ). Por lo tanto, una deflexión puede aproximarse mediante un aumento discreto en el desplazamiento angular, y el disparo de AP depende principalmente de la magnitud del desplazamiento angular.
Una desviación se definió como una combinación de desplazamiento angular hacia adelante y hacia atrás, similar a lo que ocurre cuando una presa toca un cabello. Cada medición consistió en dos deflexiones posteriores con un intervalo de 1 s entre ellas hasta un determinado desplazamiento angular θ. Si la trampa no se cerraba, seguía una fase de espera (2 minutos) para asegurar que el RP se reiniciara por completo. La espera de exactamente dos minutos, y ni más ni menos, se debe al hecho de que una serie de experimentos anteriores demostraron que a temperaturas inferiores a 30 ° C, se requieren dos desviaciones del cabello en 30-40 s para cerrar la trampa rápida y completamente.
Después de la etapa de espera, se repitió el procedimiento con desplazamientos angulares crecientes hasta que se activó el mecanismo de cierre de la trampa ( 1C y 1D). Esto ocurrió cuando se alcanzó el umbral de desplazamiento medio θ = 0,18 rad o el umbral de par medio τ = 0,8 μN · m (n = 21).
Los científicos señalan que el cierre de la trampa nunca se registró si θ estaba por debajo de 0,12 rad y τ estaba por debajo de 0,50 μN · m ( 1E ). En consecuencia, estos indicadores son el límite inferior de la desviación angular que se requiere para activar el cierre de la trampa en las condiciones de este experimento. También se pudo determinar que para que la trampa funcione, el insecto debe aplicar una fuerza (F) de 0.5 mN cerca de las puntas de los pelos o 5 mN cerca de la base.
Las mediciones de AP proporcionaron un vínculo entre la deflexión sensorial del cabello y el cierre de la trampa. Cuando dos desviaciones consecutivas estaban muy por debajo del umbral de sesgo (θ << 0,12 rad), no se observó AP. Para las amplitudes de deflexión cercanas al umbral de desplazamiento (θ <0,12 rad), se detectó un solo AP después de la segunda deflexión.
Esto indicó que ambas deflexiones del cabello contribuyeron a la RP y que el umbral de inducción de la PA solo se alcanzó en la segunda deflexión. Como era de esperar, un solo AP no fue suficiente para activar la trampa.
La suposición de que cada toque desencadena AP solo es válida si la desviación de los pelos sensoriales supera el umbral de desplazamiento. Si es así, se generaron dos AP (uno para cada deflexión del cabello) y se cerró la trampa ( 1C ).
Doble deflexión de pelos sensibles que resulta en dos AP y cierre de trampa.
Estos resultados sugieren que la deflexión rápida de los pelos sensibles aumenta la RP hasta un cierto nivel que depende de la amplitud de la deflexión angular. Los RP pueden apilarse y pueden convocar AP después de múltiples desviaciones si están por debajo del umbral de desviación. Sin embargo, la generación de un AP por toque ocurre solo si la deflexión de los pelos sensibles está por encima del umbral de deflexión.
En otras palabras, el potencial de acción se generará solo cuando el cabello sensible se haya "alterado" lo suficiente. Pero un potencial de acción de un solo disparo no es suficiente para desencadenar el cierre de la trampa. Esto encaja bien con la teoría de que los papamoscas no desperdiciarán energía en cerrar la trampa cada vez que algo o alguien les toque el pelo. De lo contrario, puede haber muchas trampas inactivas sin ningún retorno de energía (es decir, nutricional) (la presa logró escapar, la presa era demasiado pequeña o no era presa en absoluto, sino basura).
Dado que el aumento de RP con múltiples desviaciones es aditivo, sería lógico suponer que el desplazamiento sostenido (a largo plazo) de un cabello puede tener un efecto similar.
Para probar esta hipótesis, los científicos rechazaron el cabello sensible del papamoscas más allá del umbral de desplazamiento angular y lo encerraron en esa posición durante 30 segundos ( 1F ). Sin embargo, esto no activó las trampas.
El desplazamiento inicial provocó un único PA, tras lo cual la tensión volvió rápidamente a la línea de base, aunque los pelos permanecieron desviados.
Si el sesgo sostenido iba a contribuir a la RP, tendría que permanecer por encima del umbral, en cuyo caso se esperaba una serie de AP. Análisis adicionales han demostrado que la desalineación angular juega un papel importante en la RP, pero la deflexión estática del cabello no contribuye de ninguna manera.
Es curioso que en experimentos anteriores la situación fuera diferente: un solo desplazamiento de los pelos provocó el cierre de la trampa. Sin embargo, este no fue un desplazamiento único integral, sino muchos pequeños desplazamientos, ya que las oscilaciones que van acompañadas de la deflexión manual de un cabello son probablemente mayores que el umbral de desplazamiento angular.
La siguiente etapa del estudio fue crear un modelo de acumulación de carga electromecánica (ECB ), que sorprendentemente predice el cierre de trampas con un solo toque.
Basándose en los datos obtenidos durante los experimentos, los científicos desarrollaron un modelo simple para estudiar los límites del desplazamiento angular y la velocidad dentro de la cual reaccionarán las trampas.
En el modelo ECB, la deflexión mecánica da como resultado la acumulación de carga RP en función de la velocidad angular ω y el desplazamiento θ, mientras que las cargas se disipan continuamente. Si las cargas acumuladas superan un cierto umbral Q th RP , se produce un AP. Adicionalmente, se ha implementado un período refractario t RP , que representa el intervalo de tiempo requerido para que se active otro AP.
Como resultado, el modelo predijo que si las desviaciones son demasiado rápidas y / o demasiado pequeñas, es posible que se requieran más de dos de estas desviaciones para hacer que la trampa se cierre (área roja en 3A ).

Imagen # 3
Esto se debe al hecho de que una desviación no es suficiente para activar AP ( 3B), como muestran los experimentos prácticos ( 1C , centro).
De manera similar, el modelo mostró que velocidades angulares muy bajas (ω <0.04 rad / s) no pueden llenar RP.
Pero el resultado más inesperado de la simulación fue la predicción del rango de velocidades angulares intermedias (0.04 rad / s <ω <10 rad / s), en el cual una desviación es suficiente para activar dos o más AP, que son necesarios para cerrar la trampa ( 3B ).
Sin embargo, tal predicción contradice la idea de cómo funciona el mecanismo de la trampa para moscas Venus. Por tanto, los científicos decidieron comprobar si esta situación modelo se puede realizar en la práctica.
Una sola desviación de los pelos sensibles resulta en un cierre de trampa.
Sorprendentemente, el cierre de la trampa ocurrió con una sola desviación de los pelos si las velocidades angulares eran lo suficientemente bajas.
Para reducir el rango de valores en los que esto ocurre, los científicos desviaron repetidamente los mismos pelos sensoriales a diferentes velocidades angulares hasta que la trampa se cerró. Hubo un espacio de 2 minutos entre dos deflexiones consecutivas hasta que la trampa se recuperó y el RP se agotó.
El límite inferior de la velocidad angular ω requerido para cerrar la trampa en una deflexión se determinó aumentando gradualmente la velocidad angular después de cada deflexión (n = 17). La velocidad inicial estaba por debajo de 0,009 rad / s, ya que la trampa nunca se cerró a esta velocidad.
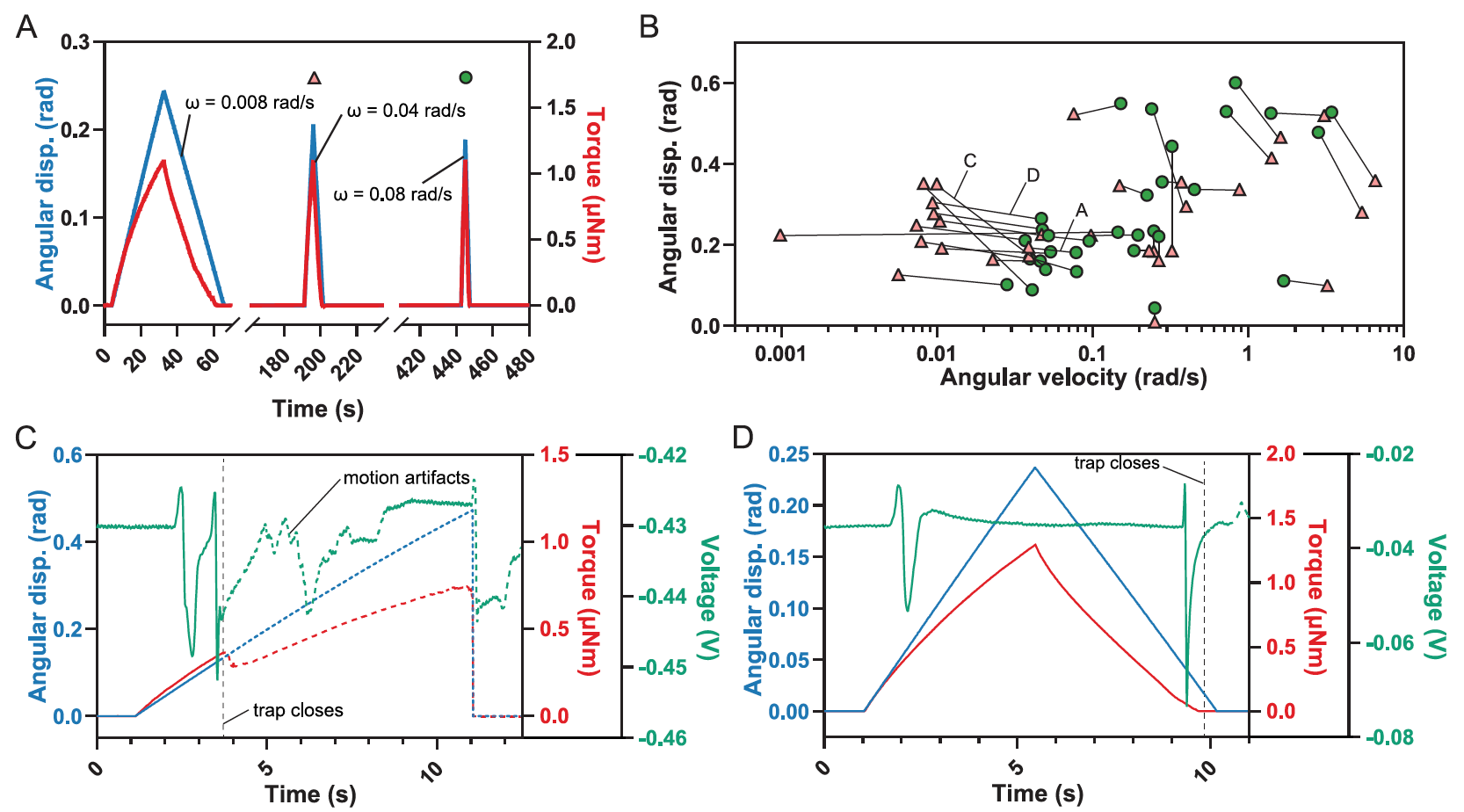
Imagen # 4 Se realizaron
posteriores deflexiones de un solo cabello con un aumento gradual de la velocidad hasta que se activó la trampa ( 4A ).
El límite superior se determinó de manera similar, comenzando con una velocidad de ω> 3 rad / s seguida de una disminución gradual (n = 9).
Además, se realizó otra serie de experimentos con una desviación (n = 5), en los que la velocidad de la sonda de fuerza se mantuvo constante, lo que llevó a una velocidad angular intermedia entre 0.2 y 0.4 rad / s, mientras que el desplazamiento angular θ aumentó gradualmente durante el tiempo de las deflexiones posteriores para obtener el límite inferior θ necesario para que la trampa funcione en condiciones de una única deflexión de los pelos.
Todas las deflexiones únicas que conducen al cierre de la trampa, junto con los estímulos anteriores para los que no se produjo el cierre de la trampa, identificaron la región donde una única deflexión provoca el cierre ( 4B ).
Una sola deflexión puede hacer que la trampa se cierre a velocidades angulares intermedias de deflexión (0,03 rad / s ≤ ω ≤ 4 rad / s). Pero la desviación sola a velocidades más bajas o más altas no será suficiente.
Los experimentos, en los que la velocidad de deflexión de los pelos era bastante baja, mostraron que la trampa se activaba tanto cuando el sensor se acercaba a la base del pelo como cuando el sensor se alejaba de él.
La desviación única de los pelos sensibles resulta en dos AP durante la flexión inicial, lo que resulta en un cierre de la trampa
Cuando se cerró la trampa durante la aproximación de la sonda, se observaron dos potenciales de acción sucesivos ( 4C ) durante la deflexión de los pelos . Cuando se cerró la trampa durante el retiro de la sonda, apareció un AP en el momento de la deflexión del cabello, y el segundo después del retiro de la sonda, cuando el cabello volvió a su posición original ( 4D ). En ambos casos, el segundo AR llevó al cierre inmediato de la trampa.
En conclusión, los científicos decidieron medir la fuerza de cierre de la trampa ( 5A ).
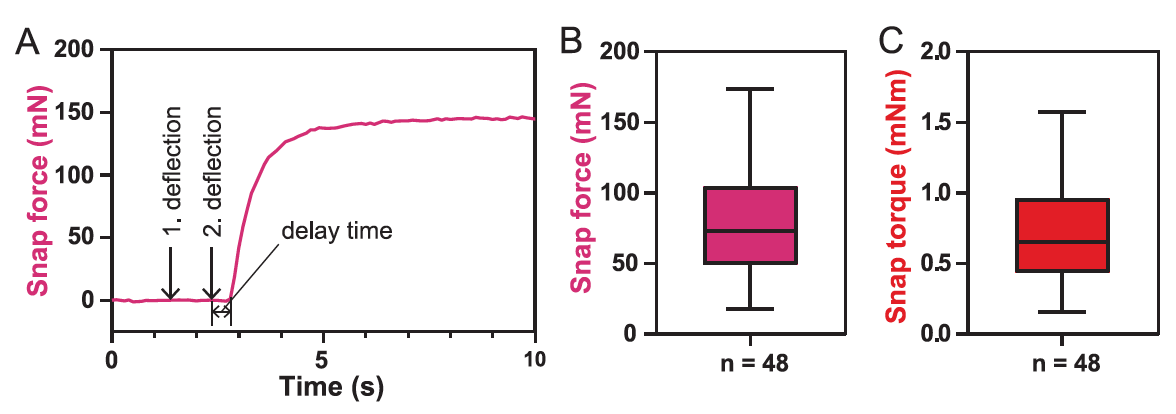
Imagen No. 5
Para la trampa 48, el valor promedio de la fuerza de agarre F de cierre fue de 73 mN ( 5V), y el rango de valores fue de 18 a 174 mN.
Estos valores son inferiores a los publicados anteriormente (140-150 mN). Hay una explicación para esto: en este caso, la fuerza se midió al comienzo del cierre de la trampa, y en observaciones anteriores, se midió la fuerza que actúa sobre los bordes de las valvas de la trampa de hojas al finalizar el cierre.
Dado que la fuerza medida depende en gran medida de la posición del transductor y también de la orientación y el tamaño de la hoja, el par de cierre τ cerca del centro de la hoja con un valor promedio de 0,65 mN · m es el mejor valor para describir la fuerza de cierre de la trampa ( 5C ). El tiempo de demora, es decir, el tiempo entre el estímulo mecánico y el inicio del cierre de la trampa, fue de 0,6 ± 0,3 s.
Para un conocimiento más detallado de los matices del estudio, le recomiendo que consulteinforme de científicos .
Epílogo
La trampa para moscas de Venus no corre como un guepardo, no tiene veneno mortal como una cobra y ciertamente no oye a su presa como un búho. Pero esta planta es de hecho uno de los mejores carnívoros del planeta, ya que su mecanismo de caza funciona como un reloj.
En este trabajo, los científicos pudieron determinar cuáles deberían ser estos o aquellos parámetros para que la trampa funcione. Al final resultó que, en la mayoría de los casos, para activar la trampa, la víctima debe cometer dos errores consecutivos: primero, tocar el cabello sensorial de la trampa para moscas; el segundo es hacerlo de nuevo.
La repetida desviación del cabello de su posición inicial conduce a la generación de un segundo potencial de acción, que inicia el proceso de cierre de la trampa. No obstante, el modelado ha demostrado que, en determinadas condiciones, un toque puede ser suficiente, pero en realidad es muy probable que esto sea poco probable.
En otras palabras, ¿puede una Venus atrapamoscas servir como cazador de mosquitos? No, porque estos chupasangres son muy pequeños y demasiado ágiles. Pero el papamoscas no se arrepiente de moscas ni caracoles.
Gracias por su atención, ¡tengan curiosidad y tengan un gran fin de semana chicos! :)
Un poco de publicidad
Gracias por estar con nosotros. ¿Te gustan nuestros artículos? ¿Quieres ver contenido más interesante? Apóyanos haciendo un pedido o recomendando a amigos, VPS en la nube para desarrolladores desde $ 4.99 , un análogo único de los servidores de nivel de entrada que hemos inventado para usted: Toda la verdad sobre VPS (KVM) E5-2697 v3 (6 núcleos) 10GB DDR4 480GB SSD 1Gbps desde $ 19 o cómo dividir el servidor correctamente? (opciones disponibles con RAID1 y RAID10, hasta 24 núcleos y hasta 40GB DDR4).
¿Dell R730xd es 2 veces más barato en el centro de datos Equinix Tier IV en Ámsterdam? ¡Solo tenemos 2 x Intel TetraDeca-Core Xeon 2x E5-2697v3 2.6GHz 14C 64GB DDR4 4x960GB SSD 1Gbps 100 TV desde $ 199 en los Países Bajos!Dell R420 - 2x E5-2430 2.2Ghz 6C 128GB DDR3 2x960GB SSD 1Gbps 100TB - ¡Desde $ 99! Lea sobre Cómo construir la infraestructura de bldg. clase con servidores Dell R730xd E5-2650 v4 a un costo de 9000 euros por un centavo?